Kyung-Ok Shin1,
Kyung-Soon Choi1,
Yong-Hwan Kim2 ![]()
For correspondence:- Yong-Hwan Kim Email: yhkim@kyonggi.ac.kr Tel:+8231249-9652
Received: 3 January 2016 Accepted: 19 May 2016 Published: 28 June 2016
Citation: Shin K, Choi K, Kim Y. In vitro antioxidative activity of moss extract, and effect of moss on serum lipid level of mice fed with high-fat diet. Trop J Pharm Res 2016; 15(6):1215-1224 doi: 10.4314/tjpr.v15i6.14
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To evaluate the potential of active compounds derived from moss in the prevention and treatment of various diseases.
Methods: Three species of moss were extracted with deionized water at 95 °C, and with 70.5 % ethanol at 85 °C. Analysis of total phenolic contents (TPC) of the extracts were performed by Folin-Ciocalteu (FC) method. The antioxidant activity of the extracts were determined using three methods, namely, by 2,2'-azino-bis(3-ethylbenzothiazoline-6-sulphonic) acid (ABTS), 1,1-diphenyl-2-picrylhydrazyl (DPPH) and ferric reducing antioxidant power (FRAP). In vivo effects were evaluated in mice fed high fat diet (HFD) supplemented with 20 % ground moss. Cholesterol levels in HFD were evaluated by o-phthalaldehyde method. Serum triglyceride levels were measured using triglyceride (TG) kit, while blood insulin level and leptin concentration were measured by enzyme-linked immunosorbent assay (ELISA) kit.
Results: The moss extracts exhibited antioxidative effects, as evidenced of . TPC of 47.20 ± 11.20 to 119.87 ± 11.51 mg GAE/mg, respectively. ABTS scavenging activity was 1078.11 ± 18.95 to 2587.33 ± 46.19 μmol Trolox/mg, DPPH scavenging activity of were 42.11 ± 8.22 to 298.78 ± 20.02 μmol Trolox/mg, and FRAP value of 393.19 ± 24.64 to 1070.14 ± 17.92 μmol Trolox/mg, respectively. Mice fed with 20 % ground moss did not show any significant effect (p < 0.05) on visceral weight and blood lipid levels of HFD, while leptin concentrations reduced significantly to 4.74 ± 0.00 and 0.20 ± 0.00 ng/dL) relative to HFD alone (26.72 ± 6.53 ng/dL).
Conclusion: Moss can potentially be used as an antioxidative ingredient, for the improvement of overall human health, suggesting that important medical benefits associated with moss consumption. However, further investigations are required to ascertain this.
Introduction
The trend of using natural ingredients for the personal health has attracted great interest of the consumer. Accordingly, a number of studies on the new natural functional resource derived from plants has been in progress [1-4]. The moss, flowerless plants collectively called bryophytes, are an important component of ecosystems at higher latitudes [5]. Moss as a plant family originated 500 million years ago and are currently represented by approximately 10,000∼16,500 species that colonize diverse habitats, ranging from high mountains to deep forests and from Antarctica to the deserts [6]. For much of human history, moss have been regarded as important natural resources, producing numerous bioactive chemicals and gene products not found in seed plants [7]. Furthermore, moss are generally regarded as safe for human consumption, based in part on their use as a staple feed by reindeer and other animals living in the tundra. While their uses often vary, moss have been used as a raw material for the production of food, clothing, dyes, medicines, models, and decorations [8]. Moss is still used in traditional medicines to alleviate the pain of burns by Gosuite Indians of Utha [9], and anticipated as antipyretic and diuretic drug already in the eleventh century in China [10]. In some western societies, moss are sometimes consumed in combination with raw fish eggs [8]. Many kinds of moss grow naturally in Korea, but there is a paucity of studies on their bioactive properties; deep-fried moss is available in Korea. Other countries, such as Denmark serve fresh reindeer moss as a dessert.
Despite their widespread use, little is known regarding the health effects of moss. In this study, we sought to examine the potentials of moss as a new biologically active resource for improved human health.
Methods
Plant materials and extraction
Three species of moss, Hypnum plumaeforme, Thuidium kanedae, and Leucobryum juniperoideum were purchased from a local market (collected from Mt. Uam, Cheongju, Korea; June, 2013). The moss were authenticated by Department of Life Science, Kyonggi University, Korea.
Moss materials were dried in a shade, ground in a mill, and stored at -18 °C in freezer (FC-A12AD, LG, Korea) until needed. Ground moss were used as additives for experimental animal feed. Moss were extracted with deionized water at 95 °C for 3 h, and with 70.5 % EtOH at 85 °C for 3 h using a heating mantle with a reflux condenser. Extracts were centrifuged at 8,000 rpm for 30 min at 4 °C using a centrifuge (SUPRA 22K, Hanil, Korea). Supernatants were evaporated under a vacuum at 45 °C using a rotary vacuum evaporator, and to produce a lyophilized powder using a freeze dryer (LYPH-LOCK 12, Labconco, USA) for analysis of antioxidant activity.
Preparation of samples for antioxidant analysis
Each lyophilized powder (1 g) were sonicated with 100 mL deionized water for for 1 h, and cooled to room temperature for 24 h. Which solutions were centrifuged at 8,000 rpm for 10 min at 4 °C and used within 24 h of preparation. Serial dilutions (1.25, 2.5, 5, 7.5, and 10 mg/mL) were then prepared using deionized water.
Determination of total phenolic content (TPC)
Total phenolic contents were determined using Folin-Ciocalteu (FC) reagent, with gallic acid as a standard [11]. FC reagent (100 μL) was added to 20 µL of diluted samples, mixed well, and incubated for 5 min at room temperature, followed by addition of 300 μL of 20 % sodium carbonate solution. Samples were then incubated for 2 h at room temperature in the dark, and the absorbance measured at 765 nm using a UV/Vis spectrophotometer (Ultrospec 3100 pro, Amersham Bio., Cambridge, UK). Results are expressed as gallic acid equivalents (mg GAE/g dry weight).
ABTS radical scavenging assay
A 2,2'-azino-bis(3-ethylbenzothiazoline-6-sulphonic) acid (ABTS) radical cation decolorization assay was performed as described by Re et al [12] with slight modifications. Briefly, ABTS solution was prepared by reacting 7 mM of aqueous ABTS solution with 2.45 mM potassium persulfate (K2S2O8). Samples were then incubated for 12 - 16 h in the dark, diluted in phosphate buffered saline (PBS, pH 7.4) to an absorbance value of 0.7 (± 0.02) at 734 nm, and equilibrated at 30 °C using a blank in PBS buffer. All samples were diluted to achieve an absorbance 20 - 80 % of the blank sample. Aliquots of diluted samples (10 μL) were mixed with 1 mL ABTS solution. The decrease in absorbance was measured over 15 min, and results expressed as µmoL Trolox/g dry weight. The decrease in absorbance was measured over 15 min, and results expressed as µmoL Trolox/g dry weight. Scavenging activity was calculated using the following equation:
Inhibition (%) = (AB - AA)/AB × 100
where, AB = absorbance of a blank sample at the beginning of the reaction (t = 0 min), and AA = absorbance of a test sample at the end of the reaction (t = 15 min).
DPPH radical scavenging assay
The radical-scavenging activity of extracts was determined using the DPPH assay as previously described [13,14], with slight modifications. In the course of the research work, DPPH solution was prepared daily in methanol (60 μM) and protected from light. Diluted samples (50 uL) were added to 2 mL of DPPH solution, mixed well, and incubated at room temperature for 30 min. Absorbance of remaining DPPH was detected at 515 nm. The blank sample contained methanol alone. Results expressed as µmoL Trolox/g dry weight. Scavenging activity was calculated using Eq 1.
Inhibition (%) = (AB - AA)/AB × 100 ………… (1)
where, AB = absorbance of a blank sample at the beginning of the reaction (t = 0 min), and AA = absorbance of test sample at the end of the reaction (t = 30 min).
Reducing power assay
The antioxidant capacity of moss, indicative of the reducing power, was estimated using the procedure described by Pulido et al [15]. Ferric reducing antioxidant power (FRAP) reagent (25 mL of 300 mM acetate buffer (pH 3.6), 2.5 mL of 10 mM tripyridyltriazine (TPTZ) solution in 40 mM HCl, and 2.5 mL of 20 mM FeCl3·6H2O was prepared fresh and warmed to 37 °C prior to the experiment. Deionized water (90 μL) was combined with 30 μL of sample as the reagent blank. The final dilution of sample in the reaction mixture was 1 : 34. A small aliquot of each sample was incubated at room temperature for 30 min. The absorbance of each sample was measured at 595 nm using a UV/Vis spectrophotometer, with results expressed as TEAC values.
Animal experiments
Experimental 8-week-old ICR mice were purchased from Orient Bio Co., Ltd, with 5 mice used per experimental group. Upon arrival at the research facility, the mice were acclimatized to solid feed (PicoLab Rodent Diet, Hwaseong, Gyeonggi, Korea), available ad libitum for 1 week, and then randomly assigned and housed with water and feed provided for 60 d. During the experimental period, mice were housed at 20 ± 2 °C, 40 - 60 % humidity and 11 ± 1 h light/dark cycle. The experimental period lasted for 8 weeks were all animal procedures (approval number: SYUIACUC 2013006) were performed in accordance with the guidelines of the Adventist University Animal Experiment Ethics Committee (IACUC Institutional Animal Care and Use Committees). Commercially available solid mouse feed was obtained in powder form and used to formulate the feed provided to the experimental animals. The composition of feed for the animal experimental diets are shown in . The composition of feed for the animal experiments included 33 to 60 % carbohydrates (starch + sucrose + glucose + fructose + lactose), 21 % proteins, and 13 to 20 % lipids (corn oil) for control to high fat diet. And were added to 1 % and 3 % vitamins and minerals, fibers were also added to 2 %. Supplemented feed were prepared to 100 % by adding 20 % ground moss.
The weights of experimental animals organs were determined in each group after abstinence from feed for 12 h prior to sacrifice. The animals were slightly anesthetized with ethyl ether, organs were removed after abdominal incision. Exposed organs were washed with cold physiological saline and weighed after removed connective tissue. Blood was collected from the heart using a syringe, incubated at 4 °C for ~1 h, and the serum separated by centrifuged at 3,000 rpm for 15 min at 5 °C using a centrifuge (A32010, Gyrozen, Korea). Aliquots (100 μL) of serum were placed into microtubes, which were stored at -70 °C in a deep freezer (MDF-192, Sanyo Electric Biomedical Co., Ltd. Japan) until needed.
Analysis of blood lipid content
Serum cholesterol was measured using the o-phthalaldehyde method, as described previously [16,17]. Samples were separated by mixing 0.1 mL of each sample with 0.3 mL of 33 % KOH solution and 0.3 mL of 95 % ethanol. The resulting mixture was heated in a 60 °C water bath for 15 min and subsequently cooled. Following the addition of 5.0 mL hexane was added, and 3.0 mL deionized water was added and mixed well for 1 min. The withdrawn hexane layer (1.0 mL) was subsequently concentrated and dried under nitrogen. After 2.0 mL of o-phthalaldehyde reagent and mixed with 1.0 mL of concentrated sulfuric acid, and incubated for 10 min at room temperature. Absorbance was measured at 550 nm using a UV/Vis spectrophotometer (X-ma 6300, Human Corporation, Korea) within 90 min of the addition of the sulfuric acid, and the cholesterol content quantified using a standard calibration curve.
HDL-cholesterol was assessed using the method of Cho and Choi [16]. A 0.3 mL of serum was placed in a test tube, along with 0.3 mL of precipitation reagent. After incubated for 10 min at room temperature, the tube were centrifuged at 700 × g for 10 min. Then 3.0 mL of HDL chromogenic reagent were added into 3 tubes containing either 50 μL of supernatant, 50 μL of standard solution (100 mg/dL), 50 μL deionized water as a blank, followed by heating in a water bath for 5 min at 37 °C. A blank sample was prepared using identical components, without the addition of the supernatant or standard solution. The absorbance was measured at 555 nm using a UV/Vis spectrophotometer and HDL-cholesterol content was quantified relative to the blank control.
LDL-cholesterol was assessed using the method of Cho and Choi [16]. A 0.1 mL serum and 0.1 mL standard serum were placed in a test tube, combined with 4.0 mL BLF kit reagents, vortexed for 5 sec, and then incubated for 25 min at room temperature (25 ± 3 °C). LDL-cholesterol content was then quantified by measuring absorbance at 650 nm using a UV/Vis spectrophotometer within 10 min, using deionized water as a control.
Serum triglyceride levels were measured as described by Cho and Choi [16], using a triglyceride (TG) kit (Sigma Co., St. Louis, MO, USA). Briefly, 1.0 mL TG kit reagent was added into 3 tubes containing either 10 µL of serum, 10 µL of standard solution (300 mg/dL), 10 µL of deionized water as a blank, followed by heating in a water bath for 5 min at 37 °C. TG content was assessed by measuring the absorbance at 540 nm using a UV/Vis spectrophotometer and quantified relative to the blank control.
Blood insulin level was determined by an enzyme-linked immunosorbent assay (ELISA) kit (Linco, USA), which were applied to plasma obtained on the last day of the experiment and analyzed by analytical equipment (Molecular Devices, USA). Serum concentrations of leptin were determined by ELISA kit. Assays were conducted in accordance with the manufacturer’s protocol (R&D Systems Inc., Minneapolis, MN, USA).
Statistical analysis
Each experiment was performed in triplicate. The data are reported as mean ± SD and were analyzed by SPSS (version 18.0; SPSS Inc., Chicago, IL, USA). Comparison of the averages in each group was conducted with one-way ANOVA, and analysis of significant differences between groups was conducted using Duncan's multiple tests. P < 0.05 was considered statistically significant.
Results
Total phenolic content (TPC)
TPC of deionized water extracts, and 70.5 % EtOH extracts of moss are shown in . TPC of H. plumaeforme, T. kanedae, and L. juniperoideum ranged from 47.20 ± 11.20 to 119.87 ± 11.51 mg GAE/mg in the extracts. The highest TPC (119.87 ± 11.51 mg GAE/mg) was observed for L. juniperoideum extracted with deionized water, while the lowest was T. kanedae extracted with 70.5 % ethanol.
ABTS radical scavenging activity
ABTS radical scavenging activities of deionized water extracts, and 70.5 % EtOH extracts of moss are shown in . ABTS activities of H. plumaeforme, T. kanedae, and L. juniperoideum ranged from 1078.11 ± 18.95 to 2587.33 ± 46.19 μmol Trolox/mg in the extracts. The highest ABTS radical scavenging activity (2587.33 ± 46.19 μmol Trolox/mg) was observed for H. plumaeforme extracted with deionized water, while the lowest was T. kanedae extracted with 70.5 % ethanol.
DPPH radical scavenging activity
DPPH radical scavenging activities of deionized water extracts, and 70.5 % EtOH extracts of moss are shown in . DPPH activities of H. plumaeforme, T. kanedae, and L. juniperoideum ranged from 42.11 ± 8.22 to 298.78 ± 20.02 µmol Trolox/mg in the extracts. The highest DPPH radical scavenging activity (298.78 ± 20.02 µmol Trolox/mg) was observed for H. plumaeforme extracted with deionized water, while the lowest was T. kanedae extracted with 70.5 % ethanol.
Reducing power
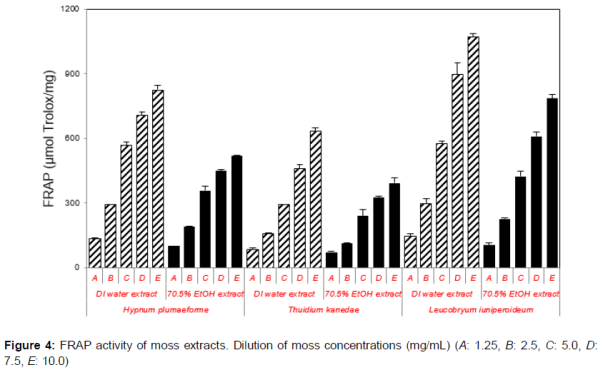
FRAP values of deionized water extracts, and 70.5 % EtOH extracts of moss are shown in . FRAP values of H. plumaeforme, T. kanedae, and L. juniperoideum ranged from 393.19 ± 24.64 to 1070.14 ± 17.92 µmol Trolox/mg in the extracts. The highest FRAP value (1070.14 ± 17.92 µmol Trolox/mg) was observed for L. juniperoideum extracted with deionized water, while the lowest was T. kanedae extracted with 70.5 % ethanol.
Organ weight and serum lipid level in the blood
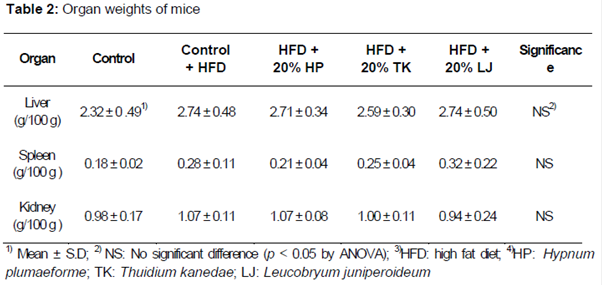
Organ weights of mice fed normal and high fat diets (HFD) supplemented with various moss are shown in . No significant differences in relative liver weight were observed between groups. Average liver weights of animals fed a HFD alone or HFD supplemented with H. plumaeforme, T. kanedae, and L. juniperoideum were 2.74 ± 0.4, 2.71 ± 0.34, 2.59 ± 0.30, and 2.74 ± 0.50 g/100 g body weight, respectively. Similarly, no significant differences were observed in the weights of other visceral organs between treatment groups.
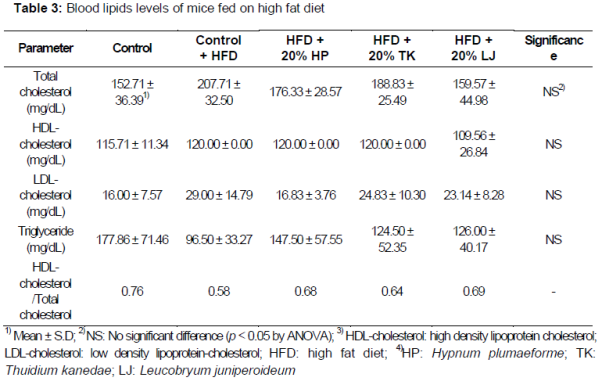
Blood lipid contents for each group are shown in . Total blood cholesterol levels of mice fed a HFD supplemented with L. juniperoideum were significantly lower (159.57 ± 44.98 mg/dL) than that of other groups. While animals fed a HFD alone exhibited the highest LDL-cholesterol levels (29.00 ± 14.79 mg/dL), these levels were not statistically different from other groups because the feed was not high in fat. Similarly, experimental animals fed a HFD supplemented with H. plumaeforme, T. kanedae, or L. juniperoideum exhibited lower plasma TG levels relative to the control group, but these differences were not statistically significant. No significant differences in HDL-cholesterol levels were observed between the groups.
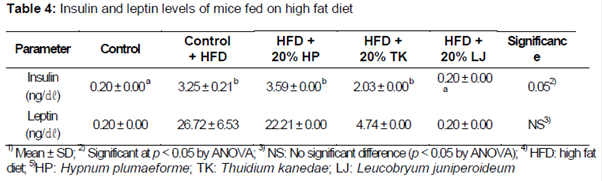
Comparison of insulin and leptin levels are shown in . In the case of blood insulin concentrations, groups fed a HFD supplemented with T. kanedae (2.03 ± 0.00 ng/dL) or L. juniperoideum exhibited significantly lower insulin concentrations relative to mice receiving a HFD alone (p < 0.05). Similarly, animals fed a HFD supplemented with T. kanedae or L. juniperoideum exhibited lower blood leptin concentrations (4.74 ± 0.00 and 0.20 ± 0.00 ng/dL, respectively) relative to mice receiving a HFD alone (26.72 ± 6.53 ng/dL).
Discussion
We evaluated the antioxidant activities by measuring total phenolic content, ABTS scavenging activity, DPPH scavenging activity, and reducing power via FRAP assay, using moss extracts obtained using both deionized water and 70.5 % EtOH.
All of the moss species described here exhibited high degrees of antioxidative activity, an important finding given the lack of studies evaluating the antioxidative properties of moss extracts. While direct comparisons of these results to other mosses are not currently possible, a variety of studies evaluating the effects of various plant extracts [18-22] are available. The polyphenol contents of teas prepared from domestic esculent plants, such as black tea, ginseng tea, and green tea have been reported to be 101.51, 28.30, and 94.90 μg/mg, respectively [19]. An alternative study evaluating kohlrabi and radish sprouts exhibited higher anti-oxidative activities, as determined by both DPPH and ABST assays [21]. Ethanol extracts of five edible spring flowers in Korea such as Forsythia koreana Nakai, Rhododendron mucronulatum Turcz., Prunus avium, Magnolia lilyflora, and Magnolia denudate have been reported high anti-oxidative activity, which are 12.8 to 46.1 mM Trolox/mg ABTS activity, 1.2 to 43.3 mM Trolox/mg DPPH activity, and 4.1 to 35.4 mM Trolox/mg FRAP activity, respectively [20]. Similarly, an extract derived from Perilla frutescens sprouts was shown to exhibit potent anti-oxidative properties, including the ability to effectively scavenge DPPH and ABTS radicals, anti-inflammatory activity, and antiemetic effects [22].
Supplementation of animal feed with moss extracts was unable to overcome many of the effects of a HFD, with no significant differences observed in the weights of visceral organs. While the group fed a HFD supplemented with L. juniperoideum did exhibit lower levels of total cholesterol in the blood, no significant differences in HDL or LDL cholesterol were observed between groups, despite modest increases in LDL levels observed in HFD controls.
Previous studies examining cholesterol levels in the blood of white rats revealed significant variability in HDL levels ranging from 20 - 92 mg/dL, and neutral fat levels ranging from 27 -108 mg/dL [23], though a subsequent analysis did confirm these results, observing normal HDL-cholesterol levels of 68.94 mg/dL (1.78 mmol/L), and neutral fat level observed only 78.77 mg/dL (0.9 mmol/L) under normal conditions, suggesting a much greater range than initially thought [24]. Other lipid components were also highly variable, with LDL-cholesterol levels ranging from 10.47 to 82.7 mg/dL [25]. Choi et al [26] reported that experimental animals fed an oil extracted from the seed of Prunus sargentii R. exhibited the highest levels of HDL-cholesterol in blood, as well as lower concentrations of LDL-cholesterol. The data presented here showed lower concentrations of both total and HDL-cholesterol in all animal groups tested, while LDL-cholesterol levels were consistent with previous publications [23,25].
Insulin levels in animals fed a HFD alone were significantly lower than those observed in animals fed a HFD supplemented with T. kanedae or L. juniperoideum (p < 0.05). Insulin resistance, which is defined as a state where insulin continues to be secreted from β-cells in the pancreas, but its physiological function is reduced, has been reported in obese patients, as well as those with abnormal glucose tolerance brought about by a diet with high fat content [27]. The eating of adequate amounts of vegetables with high antioxidant activity has been shown to reduce oxidative stress and help prevent various diseases, including insulin resistance [28] and identifying vegetables or other food that could reduce oxidative stress, and prevent insulin resistance is important. For example, consumption of Suaeda japonica and Spergularia marina has been shown to suppress insulin resistance and lower triglyceride and total cholesterol levels [29]. Controlling lipogenesis and suppressing the delivery of fats to adipocytes, while increasing the consumption of fats stored in adipocytes are important for both preventing and reducing obesity [30]. In a previous study, animals fed a HFD for 2 weeks exhibited blood leptin concentrations 2.3 fold higher than that of animals fed a normal diet, leading the authors to conclude that a HFD increases the concentration of leptin in the blood [31]. As increased leptin levels due to a HFD are directly related to the accumulation of body fat and weight gain, leptin levels cannot be lowered to the level observed in the control group [31]. Here, we found that blood leptin concentrations were elevated in animals fed a HFD, consistent with previous findings, but that these levels could be reduced by supplementing with T. kanedae and L. juniperoideum.
Further insight into the mechanisms underlying increased blood leptin concentrations can be seen in a study by Kim et al [30]. In this study, supplementation with pine nut oil was shown to suppress the appetite by increasing secretion of cholecystokinin and glucagon like peptide-1 (GLP-1), resulting in an increase in leptin production. Pine nut oil was chosen due to its high percentage of unsaturated fatty acids, with a strong potential for anti-obesity applications. Another study, which focused on the relationship between insulin and leptin, reported that insulin can increase leptin levels in the blood when injected into the human body [31], suggesting that insulin may be an important factor regulating leptin production by promoting its synthesis in humans and rodents.
Conclusion
The findings of this study suggest that moss possesses strong antioxidative activity, which can potentially be used to control obesity, diabetes and improve health generally.
Declarations
Acknowledgement
References
Archives


News Updates